Table of Contents

Chapter 15
Body Fluids and Circulation
15.1 Blood
15.2 Lymph (Tissue Fluid)
15.3 Circulatory Pathways
15.4 Double Circulation
15.5 Regulation of Cardiac Activity
15.6 Disorders of Circulatory System
You have learnt that all living cells have to be provided with nutrients, O2 and other essential substances. Also, the waste or harmful substances produced, have to be removed continuously for healthy functioning of tissues. It is therefore, essential to have efficient mechanisms for the movement of these substances to the cells and from the cells. Different groups of animals have evolved different methods for this transport. Simple organisms like sponges and coelenterates circulate water from their surroundings through their body cavities to facilitate the cells to exchange these substances. More complex organisms use special fluids within their bodies to transport such materials. Blood is the most commonly used body fluid by most of the higher organisms including humans for this purpose. Another body fluid, lymph, also helps in the transport of certain substances. In this chapter, you will learn about the composition and properties of blood and lymph (tissue fluid) and the mechanism of circulation of blood is also explained herein.
15.1 Blood
Blood is a special connective tissue consisting of a fluid matrix, plasma, and formed elements.
15.1.1 Plasma
Plasma is a straw coloured, viscous fluid constituting nearly 55 per cent of the blood. 90-92 per cent of plasma is water and proteins contribute 6-8 per cent of it. Fibrinogen, globulins and albumins are the major proteins. Fibrinogens are needed for clotting or coagulation of blood. Globulins primarly are involved in defense mechanisms of the body and the albumins help in osmotic balance. Plasma also contains small amounts of minerals like Na+, Ca++, Mg++, HCO3–, Cl–, etc. Glucose, amino acids, lipids, etc., are also present in the plasma as they are always in transit in the body. Factors for coagulation or clotting of blood are also present in the plasma in an inactive form. Plasma without the clotting factors is called serum.
15.1.2 Formed Elements
Erythrocytes, leucocytes and platelets are collectively called formed elements (Figure 15.1) and they constitute nearly 45 per cent of the blood.
Erythrocytes or red blood cells (RBC) are the most abundant of all the cells in blood. A healthy adult man has, on an average, 5 millions to 5.5 millions of RBCs mm–3 of blood. RBCs are formed in the red bone marrow in the adults. RBCs are devoid of nucleus in most of the mammals and are biconcave in shape. They have a red coloured, iron containing complex protein called haemoglobin, hence the colour and name of these cells. A healthy individual has 12-16 gms of haemoglobin in every 100 ml of blood. These molecules play a significant role in transport of respiratory gases. RBCs have an average life span of 120 days after which they are destroyed in the spleen (graveyard of RBCs).
Leucocytes are also known as white blood cells (WBC) as they are colourless due to the lack of haemoglobin. They are nucleated and are relatively lesser in number which averages 6000-8000 mm–3 of blood. Leucocytes are generally short lived. We have two main categories of WBCs – granulocytes and agranulocytes. Neutrophils, eosinophils and basophils are different types of granulocytes, while lymphocytes and monocytes are the agranulocytes. Neutrophils are the most abundant cells (60-65 per cent) of the total WBCs and basophils are the least (0.5-1 per cent) among them. Neutrophils and monocytes (6-8 per cent) are phagocytic cells which destroy foreign organisms entering the body. Basophils secrete histamine, serotonin, heparin, etc., and are involved in inflammatory reactions. Eosinophils (2-3 per cent) resist infections and are also associated with allergic reactions. Lymphocytes (20-25 per cent) are of two major types – ‘B’ and ‘T’ forms. Both B and T lymphocytes are responsible for immune responses of the body.
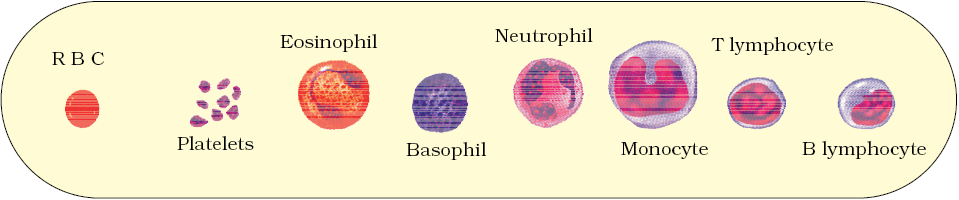
Figure 15.1 Diagrammatic representation of formed elements in blood
Platelets also called thrombocytes, are cell fragments produced from megakaryocytes (special cells in the bone marrow). Blood normally contains 1,500,00-3,500,00 platelets mm–3. Platelets can release a variety of substances most of which are involved in the coagulation or clotting of blood. A reduction in their number can lead to clotting disorders which will lead to excessive loss of blood from the body.
15.1.3 Blood Groups
As you know, blood of human beings differ in certain aspects though it appears to be similar. Various types of grouping of blood has been done. Two such groupings – the ABO and Rh – are widely used all over the world.
15.1.3.1 ABO grouping
ABO grouping is based on the presence or absence of two surface antigens (chemicals that can induce immune response) on the RBCs namely A and B. Similarly, the plasma of different individuals contain two natural antibodies (proteins produced in response to antigens). The distribution of antigens and antibodies in the four groups of blood, A, B, AB and O are given in Table 15.1. You probably know that during blood transfusion, any blood cannot be used; the blood of a donor has to be carefully matched with the blood of a recipient before any blood transfusion to avoid severe problems of clumping (destruction of RBC). The donor’s compatibility is also shown in the Table 15.1.
Table 15.1 Blood Groups and Donor Compatibility
| Blood Group | Antigens on RBCs | Antibodies in Plasma | Donor’s Group |
| A | A | anti-B | A,O |
| B | B | anti-A | B,O |
| AB | A,B | nil | AB, A, B, O |
| O | nil | anti-A,B | O |
From the above mentioned table it is evident that group ‘O’ blood can be donated to persons with any other blood group and hence ‘O’ group individuals are called ‘universal donors’. Persons with ‘AB’ group can accept blood from persons with AB as well as the other groups of blood. Therefore, such persons are called ‘universal recipients’.
15.1.3.2 Rh grouping
Another antigen, the Rh antigen similar to one present in Rhesus monkeys (hence Rh), is also observed on the surface of RBCs of majority (nearly 80 per cent) of humans. Such individuals are called Rh positive (Rh+ve) and those in whom this antigen is absent are called Rh negative (Rh-ve). An Rh-ve person, if exposed to Rh+ve blood, will form specific antibodies against the Rh antigens. Therefore, Rh group should also be matched before transfusions. A special case of Rh incompatibility (mismatching) has been observed between the Rh-ve blood of a pregnant mother with Rh+ve blood of the foetus. Rh antigens of the foetus do not get exposed to the Rh-ve blood of the mother in the first pregnancy as the two bloods are well separated by the placenta. However, during the delivery of the first child, there is a possibility of exposure of the maternal blood to small amounts of the Rh+ve blood from the foetus. In such cases, the mother starts preparing antibodies against Rh antigen in her blood. In case of her subsequent pregnancies, the Rh antibodies from the mother (Rh-ve) can leak into the blood of the foetus (Rh+ve) and destroy the foetal RBCs. This could be fatal to the foetus or could cause severe anaemia and jaundice to the baby. This condition is called erythroblastosis foetalis. This can be avoided by administering anti-Rh antibodies to the mother immediately after the delivery of the first child.
15.1.4 Coagulation of Blood
You know that when you cut your finger or hurt yourself, your wound does not continue to bleed for a long time; usually the blood stops flowing after sometime. Do you know why? Blood exhibits coagulation or clotting in response to an injury or trauma. This is a mechanism to prevent excessive loss of blood from the body. You would have observed a dark reddish brown scum formed at the site of a cut or an injury over a period of time. It is a clot or coagulam formed mainly of a network of threads called fibrins in which dead and damaged formed elements of blood are trapped. Fibrins are formed by the conversion of inactive fibrinogens in the plasma by the enzyme thrombin. Thrombins, in turn are formed from another inactive substance present in the plasma called prothrombin. An enzyme complex, thrombokinase, is required for the above reaction. This complex is formed by a series of linked enzymic reactions (cascade process) involving a number of factors present in the plasma in an inactive state. An injury or a trauma stimulates the platelets in the blood to release certain factors which activate the mechanism of coagulation. Certain factors released by the tissues at the site of injury also can initiate coagulation. Calcium ions play a very important role in clotting.
15.2 Lymph (Tissue Fluid)
As the blood passes through the capillaries in tissues, some water along with many small water soluble substances move out into the spaces between the cells of tissues leaving the larger proteins and most of the formed elements in the blood vessels. This fluid released out is called the interstitial fluid or tissue fluid. It has the same mineral distribution as that in plasma. Exchange of nutrients, gases, etc., between the blood and the cells always occur through this fluid. An elaborate network of vessels called the lymphatic system collects this fluid and drains it back to the major veins. The fluid present in the lymphatic system is called the lymph. Lymph is a colourless fluid containing specialised lymphocytes which are responsible for the immune responses of the body. Lymph is also an important carrier for nutrients, hormones, etc. Fats are absorbed through lymph in the lacteals present in the intestinal villi.
15.3 Circulatory Pathways
The circulatory patterns are of two types – open or closed. Open circulatory system is present in arthropods and molluscs in which blood pumped by the heart passes through large vessels into open spaces or body cavities called sinuses. Annelids and chordates have a closed circulatory system in which the blood pumped by the heart is always circulated through a closed network of blood vessels. This pattern is considered to be more advantageous as the flow of fluid can be more precisely regulated.
All vertebrates possess a muscular chambered heart. Fishes have a 2-chambered heart with an atrium and a ventricle. Amphibians and the reptiles (except crocodiles) have a 3-chambered heart with two atria and a single ventricle, whereas crocodiles, birds and mammals possess a 4-chambered heart with two atria and two ventricles. In fishes the heart pumps out deoxygenated blood which is oxygenated by the gills and supplied to the body parts from where deoxygenated blood is returned to the heart (single circulation). In amphibians and reptiles, the left atrium receives oxygenated blood from the gills/lungs/skin and the right atrium gets the deoxygenated blood from other body parts. However, they get mixed up in the single ventricle which pumps out mixed blood (incomplete double circulation). In birds and mammals, oxygenated and deoxygenated blood received by the left and right atria respectively passes on to the ventricles of the same sides. The ventricles pump it out without any mixing up, i.e., two separate circulatory pathways are present in these organisms, hence, these animals have double circulation. Let us study the human circulatory system.
15.3.1 Human Circulatory System
Human circulatory system, also called the blood vascular system consists of a muscular chambered heart, a network of closed branching blood vessels and blood, the fluid which is circulated.
Heart, the mesodermally derived organ, is situated in the thoracic cavity, in between the two lungs, slightly tilted to the left. It has the size of a clenched fist. It is protected by a double walled membranous bag, pericardium, enclosing the pericardial fluid. Our heart has four chambers, two relatively small upper chambers called atria and two larger lower chambers called ventricles. A thin, muscular wall called the inter-atrial septum separates the right and the left atria, whereas a thick-walled, the inter-ventricular septum, separates the left and the right ventricles (Figure 15.2). The atrium and the ventricle of the same side are also separated by a thick fibrous tissue called the atrio-ventricular septum. However, each of these septa are provided with an opening through which the two chambers of the same side are connected. The opening between the right atrium and the right ventricle is guarded by a valve formed of three muscular flaps or cusps, the tricuspid valve, whereas a bicuspid or mitral valve guards the opening between the left atrium and the left ventricle. The openings of the right and the left ventricles into the pulmonary artery and the aorta respectively are provided with the semilunar valves. The valves in the heart allows the flow of blood only in one direction, i.e., from the atria to the ventricles and from the ventricles to the pulmonary artery or aorta. These valves prevent any backward flow.
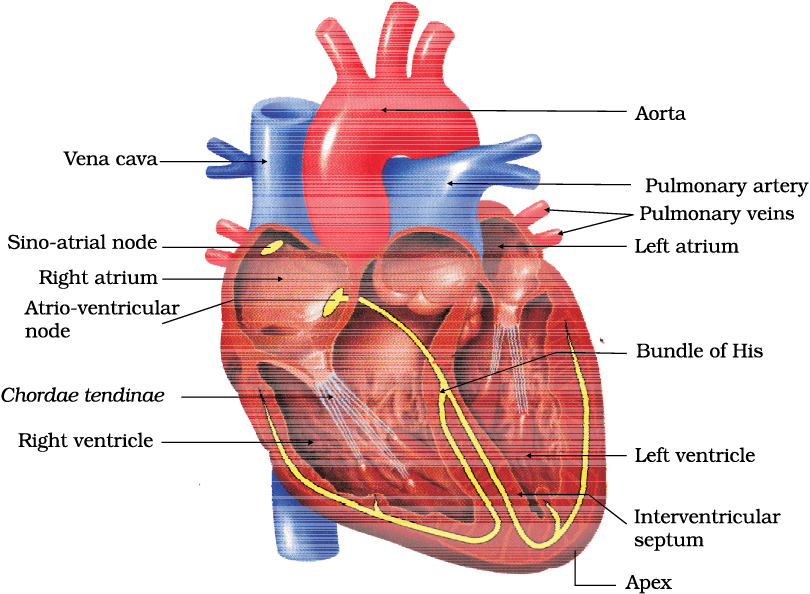
Figure 15.2 Section of a human heart
The entire heart is made of cardiac muscles. The walls of ventricles are much thicker than that of the atria. A specialised cardiac musculature called the nodal tissue is also distributed in the heart (Figure 15.2). A patch of this tissue is present in the right upper corner of the right atrium called the sino-atrial node (SAN). Another mass of this tissue is seen in the lower left corner of the right atrium close to the atrio-ventricular septum called the atrio-ventricular node (AVN). A bundle of nodal fibres, atrio-ventricular bundle (AV bundle) continues from the AVN which passes through the atrio-ventricular septa to emerge on the top of the inter-ventricular septum and immediately divides into a right and left bundle. These branches give rise to minute fibres throughout the ventricular musculature of the respective sides and are called purkinje fibres. The nodal musculature has the ability to generate action potentials without any external stimuli, i.e., it is autoexcitable. However, the number of action potentials that could be generated in a minute vary at different parts of the nodal system. The SAN can generate the maximum number of action potentials, i.e., 70-75 min–1, and is responsible for initiating and maintaining the rhythmic contractile activity of the heart. Therefore, it is called the pacemaker. Our heart normally beats 70-75 times in a minute (average 72 beats min–1).
15.3.2 Cardiac Cycle
How does the heart function? Let us take a look. To begin with, all the four chambers of heart are in a relaxed state, i.e., they are in joint diastole. As the tricuspid and bicuspid valves are open, blood from the pulmonary veins and vena cava flows into the left and the right ventricle respectively through the left and right atria. The semilunar valves are closed at this stage. The SAN now generates an action potential which stimulates both the atria to undergo a simultaneous contraction – the atrial systole. This increases the flow of blood into the ventricles by about 30 per cent. The action potential is conducted to the ventricular side by the AVN and AV bundle from where the bundle of His transmits it through the entire ventricular musculature. This causes the ventricular muscles to contract, (ventricular systole), the atria undergoes relaxation (diastole), coinciding with the ventricular systole. Ventricular systole increases the ventricular pressure causing the closure of tricuspid and bicuspid valves due to attempted backflow of blood into the atria. As the ventricular pressure increases further, the semilunar valves guarding the pulmonary artery (right side) and the aorta (left side) are forced open, allowing the blood in the ventricles to flow through these vessels into the circulatory pathways. The ventricles now relax (ventricular diastole) and the ventricular pressure falls causing the closure of semilunar valves which prevents the backflow of blood into the ventricles. As the ventricular pressure declines further, the tricuspid and bicuspid valves are pushed open by the pressure in the atria exerted by the blood which was being emptied into them by the veins. The blood now once again moves freely to the ventricles. The ventricles and atria are now again in a relaxed (joint diastole) state, as earlier. Soon the SAN generates a new action potential and the events described above are repeated in that sequence and the process continues.
This sequential event in the heart which is cyclically repeated is called the cardiac cycle and it consists of systole and diastole of both the atria and ventricles. As mentioned earlier, the heart beats 72 times per minute, i.e., that many cardiac cycles are performed per minute. From this it could be deduced that the duration of a cardiac cycle is 0.8 seconds. During a cardiac cycle, each ventricle pumps out approximately 70 mL of blood which is called the stroke volume. The stroke volume multiplied by the heart rate (no. of beats per min.) gives the cardiac output. Therefore, the cardiac output can be defined as the volume of blood pumped out by each ventricle per minute and averages 5000 mL or 5 litres in a healthy individual. The body has the ability to alter the stroke volume as well as the heart rate and thereby the cardiac output. For example, the cardiac output of an athlete will be much higher than that of an ordinary man.
During each cardiac cycle two prominent sounds are produced which can be easily heard through a stethoscope. The first heart sound (lub) is associated with the closure of the tricuspid and bicuspid valves whereas the second heart sound (dub) is associated with the closure of the semilunar valves. These sounds are of clinical diagnostic significance.
15.3.3 Electrocardiograph (ECG)
You are probably familiar with this scene from a typical hospital television show: A patient is hooked up to a monitoring machine that shows voltage traces on a screen and makes the sound “... pip... pip... pip..... peeeeeeeeeeeeeeeeeeeeee” as the patient goes into cardiac arrest. This type of machine (electro-cardiograph) is used to obtain an electrocardiogram (ECG). ECG is a graphical representation of the electrical activity of the heart during a cardiac cycle. To obtain a standard ECG (as shown in the Figure 15.3), a patient is connected to the machine with three electrical leads (one to each wrist and to the left ankle) that continuously monitor the heart activity. For a detailed evaluation of the heart’s function, multiple leads are attached to the chest region. Here, we will talk only about a standard ECG.
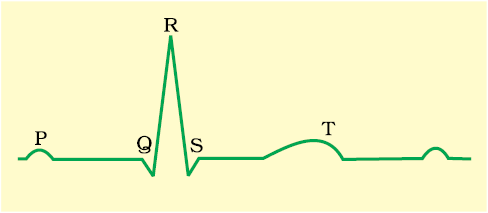
Figure 15.3 Diagrammatic presentation of a standard ECG
Each peak in the ECG is identified with a letter from P to T that corresponds to a specific electrical activity of the heart.
The P-wave represents the electrical excitation (or depolarisation) of the atria, which leads to the contraction of both the atria.
The QRS complex represents the depolarisation of the ventricles, which initiates the ventricular contraction. The contraction starts shortly after Q and marks the beginning of the systole.
The T-wave represents the return of the ventricles from excited to normal state (repolarisation). The end of the T-wave marks the end of systole.
Obviously, by counting the number of QRS complexes that occur in a given time period, one can determine the heart beat rate of an individual. Since the ECGs obtained from different individuals have roughly the same shape for a given lead configuration, any deviation from this shape indicates a possible abnormality or disease. Hence, it is of a great clinical significance.
15.4 Double Circulation
The blood flows strictly by a fixed route through Blood Vessels—the arteries and veins. Basically, each artery and vein consists of three layers: an inner lining of squamous endothelium, the tunica intima, a middle layer of smooth muscle and elastic fibres, the tunica media, and an external layer of fibrous connective tissue with collagen fibres, the tunica externa. The tunica media is comparatively thin in the veins (Figure 15.4).
As mentioned earlier, the blood pumped by the right ventricle enters the pulmonary artery, whereas the left ventricle pumps blood into the aorta. The deoxygenated blood pumped into the pulmonary artery is passed on to the lungs from where the oxygenated blood is carried by the pulmonary veins into the left atrium. This pathway constitutes the pulmonary circulation. The oxygenated blood entering the aorta is carried by a network of arteries, arterioles and capillaries to the tissues from where the deoxygenated blood is collected by a system of venules, veins and vena cava and emptied into the right atrium. This is the systemic circulation (Figure 15.4). The systemic circulation provides nutrients, O2 and other essential substances to the tissues and takes CO2 and other harmful substances away for elimination. A unique vascular connection exists between the digestive tract and liver called hepatic portal system. The hepatic portal vein carries blood from intestine to the liver before it is delivered to the systemic circulation. A special coronary system of blood vessels is present in our body exclusively for the circulation of blood to and from the cardiac musculature.
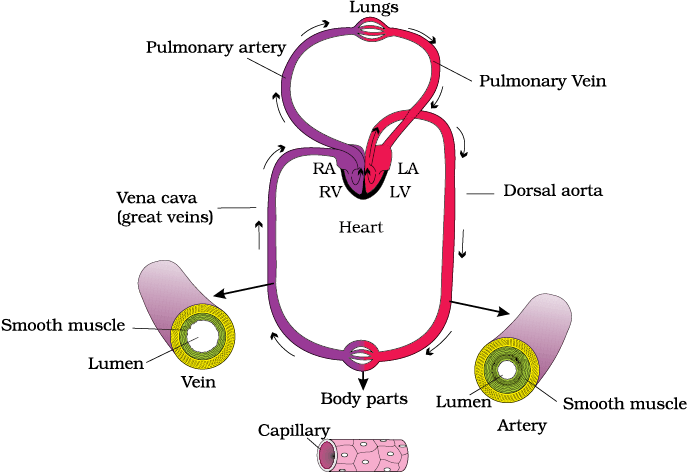
Figure 15.4 Schematic plan of blood circulation in human
15.5 Regulation of Cardiac Activity
Normal activities of the heart are regulated intrinsically, i.e., auto regulated by specialised muscles (nodal tissue), hence the heart is called myogenic. A special neural centre in the medulla oblangata can moderate the cardiac function through autonomic nervous system (ANS). Neural signals through the sympathetic nerves (part of ANS) can increase the rate of heart beat, the strength of ventricular contraction and thereby the cardiac output. On the other hand, parasympathetic neural signals (another component of ANS) decrease the rate of heart beat, speed of conduction of action potential and thereby the cardiac output. Adrenal medullary hormones can also increase the cardiac output.
15.6 Disorders of Circulatory System
High Blood Pressure (Hypertension): Hypertension is the term for blood pressure that is higher than normal (120/80). In this measurement 120 mm Hg (millimetres of mercury pressure) is the systolic, or pumping, pressure and 80 mm Hg is the diastolic, or resting, pressure. If repeated checks of blood pressure of an individual is 140/90 (140 over 90) or higher, it shows hypertension. High blood pressure leads to heart diseases and also affects vital organs like brain and kidney.
Coronary Artery Disease (CAD): Coronary Artery Disease, often referred to as atherosclerosis, affects the vessels that supply blood to the heart muscle. It is caused by deposits of calcium, fat, cholesterol and fibrous tissues, which makes the lumen of arteries narrower.
Angina: It is also called ‘angina pectoris’. A symptom of acute chest pain appears when no enough oxygen is reaching the heart muscle. Angina can occur in men and women of any age but it is more common among the middle-aged and elderly. It occurs due to conditions that affect the blood flow.
Heart Failure: Heart failure means the state of heart when it is not pumping blood effectively enough to meet the needs of the body. It is sometimes called congestive heart failure because congestion of the lungs is one of the main symptoms of this disease. Heart failure is not the same as cardiac arrest (when the heart stops beating) or a heart attack (when the heart muscle is suddenly damaged by an inadequate blood supply)
Summary
Vertebrates circulate blood, a fluid connective tissue, in their body, to transport essential substances to the cells and to carry waste substances from there. Another fluid, lymph (tissue fluid) is also used for the transport of certain substances.
Blood comprises of a fluid matrix, plasma and formed elements. Red blood cells (RBCs, erythrocytes), white blood cells (WBCs, leucocytes) and platelets (thrombocytes) constitute the formed elements. Blood of humans are grouped into A, B, AB and O systems based on the presence or absence of two surface antigens, A, B on the RBCs. Another blood grouping is also done based on the presence or absence of another antigen called Rhesus factor (Rh) on the surface of RBCs. The spaces between cells in the tissues contain a fluid derived from blood called tissue fluid. This fluid called lymph is almost similar to blood except for the protein content and the formed elements.
All vertebrates and a few invertebrates have a closed circulatory system. Our circulatory system consists of a muscular pumping organ, heart, a network of vessels and a fluid, blood. Heart has two atria and two ventricles. Cardiac musculature is auto-excitable. Sino-atrial node (SAN) generates the maximum number of action protentials per minute (70-75/min) and therefore, it sets the pace of the activities of the heart. Hence it is called the Pacemaker. The action potential causes the atria and then the ventricles to undergo contraction (systole) followed by their relaxation (diastole). The systole forces the blood to move from the atria to the ventricles and to the pulmonary artery and the aorta. The cardiac cycle is formed by sequential events in the heart which is cyclically repeated and is called the cardiac cycle. A healthy person shows 72 such cycles per minute. About 70 mL of blood is pumped out by each ventricle during a cardiac cycle and it is called the stroke or beat volume. Volume of blood pumped out by each ventricle of heart per minute is called the cardiac output and it is equal to the product of stroke volume and heart rate (approx 5 litres). The electrical activity of the heart can be recorded from the body surface by using electrocardiograph and the recording is called electrocardiogram (ECG) which is of clinical importance.
We have a complete double circulation, i.e., two circulatory pathways, namely, pulmonary and systemic are present. The pulmonary circulation starts by the pumping of deoxygenated blood by the right ventricle which is carried to the lungs where it is oxygenated and returned to the left atrium. The systemic circulation starts with the pumping of oxygenated blood by the left ventricle to the aorta which is carried to all the body tissues and the deoxygenated blood from there is collected by the veins and returned to the right atrium. Though the heart is autoexcitable, its functions can be moderated by neural and hormonal mechanisms.
Exercises
1. Name the components of the formed elements in the blood and mention one major function of each of them.
2. What is the importance of plasma proteins?
3. Match Column I with Column II :
| Column I | Column II |
| (a) Eosinophils | (i) Coagulation |
| (b) RBC | (ii) Universal Recipient |
| (c) AB Group | (iii) Resist Infections |
| (d) Platelets | (iv) Contraction of Heart |
| (e) Systole | (v) Gas transport |
4. Why do we consider blood as a connective tissue?
5. What is the difference between lymph and blood?
6. What is meant by double circulation? What is its significance?
8. Describe the evolutionary change in the pattern of heart among the vertebrates.
9. Why do we call our heart myogenic?
10. Sino-atrial node is called the pacemaker of our heart. Why?
11. What is the significance of atrio-ventricular node and atrio-ventricular bundle in the functioning of heart?
12. Define a cardiac cycle and the cardiac output.
13. Explain heart sounds.
14. Draw a standard ECG and explain the different segments in it.