
Chapter 13
BIODIVERSITY AND CONSERVATION
13.1 Biodiversity
13.2 Biodiversity Conservation
If an alien from a distant galaxy were to visit our planet Earth, the first thing that would amaze and baffle him would most probably be the enormous diversity of life that he would encounter. Even for humans, the rich variety of living organisms with which they share this planet never ceases to astonish and fascinate us. The common man would find it hard to believe that there are more than 20,000 species of ants, 3,00,000 species of beetles, 28,000 species of fishes and nearly 20,000 species of orchids. Ecologists and evolutionary biologists have been trying to understand the significance of such diversity by asking important questions– Why are there so many species? Did such great diversity exist throughout earth’s history? How did this diversification come about? How and why is this diversity important to the biosphere? Would it function any differently if the diversity was much less? How do humans benefit from the diversity of life?
13.1 Biodiversity
In our biosphere immense diversity (or heterogeneity) exists not only at the species level but at all levels of biological organisation ranging from macromolecules within cells to biomes. Biodiversity is the term popularised by the sociobiologist Edward Wilson to describe the combined diversity at all the levels of biological organisation.
The most important of them are–
(i) Genetic diversity: A single species might show high diversity at the genetic level over its distributional range. The genetic variation shown by the medicinal plant Rauwolfia vomitoria growing in different Himalayan ranges might be in terms of the potency and concentration of the active chemical (reserpine) that the plant produces. India has more than 50,000 genetically different strains of rice, and 1,000 varieties of mango.
(ii) Species diversity: The diversity at the species level, for example, the Western Ghats have a greater amphibian species diversity than the Eastern Ghats.
(iii) Ecological diversity: At the ecosystem level, India, for instance, with its deserts, rain forests, mangroves, coral reefs, wetlands, estuaries, and alpine meadows has a greater ecosystem diversity than a Scandinavian country like Norway.
It has taken millions of years of evolution, to accumulate this rich diversity in nature, but we could lose all that wealth in less than two centuries if the present rates of species losses continue. Biodiversity and its conservation are now vital environmental issues of international concern as more and more people around the world begin to realise the critical importance of biodiversity for our survival and well- being on this planet.
13.1.1 How Many Species are there on Earth and How Many in India?
Since there are published records of all the species discovered and named, we know how many species in all have been recorded so far, but it is not easy to answer the question of how many species there are on earth. According to the International Union for Conservation of Nature and Natural Resources (IUCN) (2004), the total number of plant and animal species described so far is slightly more than 1.5 million, but we have no clear idea of how many species are yet to be discovered and described. Estimates vary widely and many of them are only educated guesses. For many taxonomic groups, species inventories are more complete in temperate than in tropical countries. Considering that an overwhelmingly large proportion of the species waiting to be discovered are in the tropics, biologists make a statistical comparison of the temperate-tropical species richness of an exhaustively studied group of insects and extrapolate this ratio to other groups of animals and plants to come up with a gross estimate of the total number of species on earth. Some extreme estimates range from 20 to 50 million, but a more conservative and scientifically sound estimate made by Robert May places the global species diversity at about 7 million.
Let us look at some interesting aspects about earth’s biodiversity based on the currently available species inventories. More than 70 per cent of all the species recorded are animals, while plants (including algae, fungi, bryophytes, gymnosperms and angiosperms) comprise no more than 22 per cent of the total. Among animals, insects are the most species-rich taxonomic group, making up more than 70 per cent of the total. That means, out of every 10 animals on this planet, 7 are insects. Again, how do we explain this enormous diversification of insects? The number of fungi species in the world is more than the combined total of the species of fishes, amphibians, reptiles and mammals. In Figure 13.1, biodiversity is depicted showing species number of major taxa.
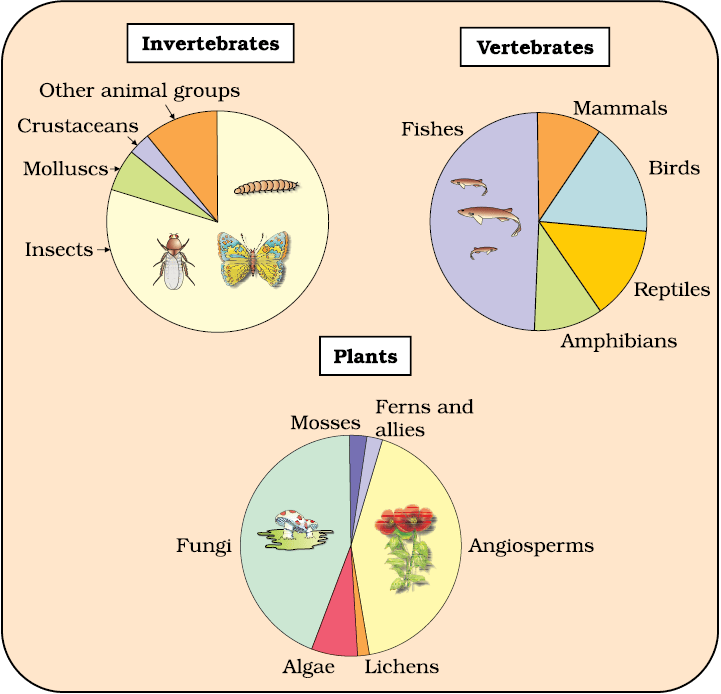
Figure 13.1 Representing global biodiversity: proportionate number of species of major taxa of plants, invertebrates and vertebrates
It should be noted that these estimates do not give any figures for prokaryotes. Biologists are not sure about how many prokaryotic species there might be. The problem is that conventional taxonomic methods are not suitable for identifying microbial species and many species are simply not culturable under laboratory conditions. If we accept biochemical or molecular criteria for delineating species for this group, then their diversity alone might run into millions.
Although India has only 2.4 per cent of the world’s land area, its share of the global species diversity is an impressive 8.1 per cent. That is what makes our country one of the 12 mega diversity countries of the world. Nearly 45,000 species of plants and twice as many of animals have been recorded from India. How many living species are actually there waiting to be discovered and named? If we accept May’s global estimates, only 22 per cent of the total species have been recorded so far. Applying this proportion to India’s diversity figures, we estimate that there are probably more than 1,00,000 plant species and more than 3,00,000 animal species yet to be discovered and described. Would we ever be able to complete the inventory of the biological wealth of our country? Consider the immense trained manpower (taxonomists) and the time required to complete the job. The situation appears more hopeless when we realise that a large fraction of these species faces the threat of becoming extinct even before we discover them. Nature’s biological library is burning even before we catalogued the titles of all the books stocked there.
13.1.2 Patterns of Biodiversity
(i) Latitudinal gradients: The diversity of plants and animals is not uniform throughout the world but shows a rather uneven distribution. For many group of animals or plants, there are interesting patterns in diversity, the most well- known being the latitudinal gradient in diversity. In general, species diversity decreases as we move away from the equator towards the poles. With very few exceptions, tropics (latitudinal range of 23.5° N to 23.5° S) harbour more species than temperate or polar areas. Colombia located near the equator has nearly 1,400 species of birds while New York at 41° N has 105 species and Greenland at 71° N only 56 species. India, with much of its land area in the tropical latitudes, has more than 1,200 species of birds. A forest in a tropical region like Equador has up to 10 times as many species of vascular plants as a forest of equal area in a temperate region like the Midwest of the USA. The largely tropical Amazonian rain forest in South America has the greatest biodiversity on earth- it is home to more than 40,000 species of plants, 3,000 of fishes, 1,300 of birds, 427 of mammals, 427 of amphibians, 378 of reptiles and of more than 1,25,000 invertebrates. Scientists estimate that in these rain forests there might be at least two million insect species waiting to be discovered and named.
What is so special about tropics that might account for their greater biological diversity? Ecologists and evolutionary biologists have proposed various hypotheses; some important ones are (a) Speciation is generally a function of time, unlike temperate regions subjected to frequent glaciations in the past, tropical latitudes have remained relatively undisturbed for millions of years and thus, had a long evolutionary time for species diversification, (b) Tropical environments, unlike temperate ones, are less seasonal, relatively more constant and predictable. Such constant environments promote niche specialisation and lead to a greater species diversity and (c) There is more solar energy available in the tropics, which contributes to higher productivity; this in turn might contribute indirectly to greater diversity.
(ii) Species-Area relationships: During his pioneering and extensive explorations in the wilderness of South American jungles, the great German naturalist and geographer Alexander von Humboldt observed that within a region species richness increased with increasing explored area, but only up to a limit. In fact, the relation between species richness and area for a wide variety of taxa (angiosperm plants, birds, bats, freshwater fishes) turns out to be a rectangular hyperbola (Figure13.2).
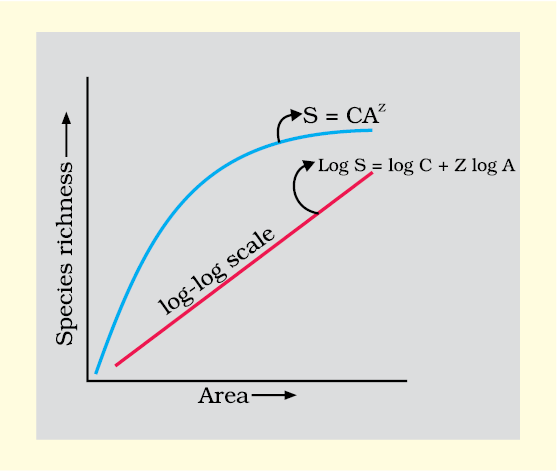
Figure 13.2 Showing species area relationship.
Note that on log scale the relationship becomes linear
On a logarithmic scale, the relationship is a straight line described by the equation
log S = log C + Z log A
where
S= Species richness A= Area
Z= slope of the line (regression coefficient)
C = Y-intercept
Ecologists have discovered that the value of z lies in the range of 0.1 to 0.2, regardless of the taxonomic group or the region (whether it is the plants in Britain, birds in California or molluscs in New York state, the slopes of the regression line are amazingly similar). But, if you analyse the species-area relationships among very large areas like the entire continents, you will find that the slope of the line to be much steeper (z values in the range of 0.6 to 1.2). For example, for frugivorous (fruit-eating) birds and mammals in the tropical forests of different continents, the slope is found to be 1.15. What do steeper slopes mean in this context?
13.1.3 The importance of Species Diversity to the Ecosystem
Does the number of species in a community really matter to the functioning of the ecosystem?This is a question for which ecologists have not been able to give a definitive answer. For many decades, ecologists believed that communities with more species, generally, tend to be more stable than those with less species. What exactly is stability for a biological community? A stable community should not show too much variation in productivity from year to year; it must be either resistant or resilient to occasional disturbances (natural or man-made), and it must also be resistant to invasions by alien species. We don’t know how these attributes are linked to species richness in a community, but David Tilman’s long-term ecosystem experiments using outdoor plots provide some tentative answers. Tilman found that plots with more species showed less year-to-year variation in total biomass. He also showed that in his experiments, increased diversity contributed to higher productivity.
Although, we may not understand completely how species richness contributes to the well-being of an ecosystem, we know enough to realise that rich biodiversity is not only essential for ecosystem health but imperative for the very survival of the human race on this planet. At a time when we are losing species at an alarming pace, one might ask– Does it really matter to us if a few species become extinct? Would Western Ghats ecosystems be less functional if one of its tree frog species is lost forever? How is our quality of life affected if, say, instead of 20,000 we have only 15,000 species of ants on earth?
There are no direct answers to such näive questions but we can develop a proper perspective through an analogy (the ‘rivet popper hypothesis’) used by Stanford ecologist Paul Ehrlich. In an airplane (ecosystem) all parts are joined together using thousands of rivets (species). If every passenger travelling in it starts popping a rivet to take home (causing a species to become extinct), it may not affect flight safety (proper functioning of the ecosystem) initially, but as more and more rivets are removed, the plane becomes dangerously weak over a period of time. Furthermore, which rivet is removed may also be critical. Loss of rivets on the wings (key species that drive major ecosystem functions) is obviously a more serious threat to flight safety than loss of a few rivets on the seats or windows inside the plane.
13.1.4 Loss of Biodiversity
While it is doubtful if any new species are being added (through speciation) into the earth’s treasury of species, there is no doubt about their continuing losses. The biological wealth of our planet has been declining rapidly and the accusing finger is clearly pointing to human activities. The colonisation of tropical Pacific Islands by humans is said to have led to the extinction of more than 2,000 species of native birds. The IUCN Red List (2004) documents the extinction of 784 species (including 338 vertebrates, 359 invertebrates and 87 plants) in the last 500 years. Some examples of recent extinctions include the dodo (Mauritius), quagga (Africa), thylacine (Australia), Steller’s Sea Cow (Russia) and three subspecies (Bali, Javan, Caspian) of tiger. The last twenty years alone have witnessed the disappearance of 27 species. Careful analysis of records shows that extinctions across taxa are not random; some groups like amphibians appear to be more vulnerable to extinction. Adding to the grim scenario of extinctions is the fact that more than 15,500 species world-wide are facing the threat of extinction. Presently, 12 per cent of all bird species, 23 per cent of all mammal species, 32 per cent of all amphibian species and 31per cent of all gymnosperm species in the world face the threat of extinction.
From a study of the history of life on earth through fossil records, we learn that large-scale loss of species like the one we are currently witnessing have also happened earlier, even before humans appeared on the scene. During the long period (> 3 billion years) since the origin and diversification of life on earth there were five episodes of mass extinction of species. How is the ‘Sixth Extinction’ presently in progress different from the previous episodes? The difference is in the rates; the current species extinction rates are estimated to be 100 to 1,000 times faster than in the pre-human times and our activities are responsible for the faster rates. Ecologists warn that if the present trends continue, nearly half of all the species on earth might be wiped out within the next 100 years.
In general, loss of biodiversity in a region may lead to (a) decline in plant production, (b) lowered resistance to environmental perturbations such as drought and (c) increased variability in certain ecosystem processes such as plant productivity, water use, and pest and disease cycles.
Causes of biodiversity losses: The accelerated rates of species extinctions that the world is facing now are largely due to human activities. There are four major causes (‘ The Evil Quartet ’ is the sobriquet used to describe them).
(i) Habitat loss and fragmentation: This is the most important cause driving animals and plants to extinction. The most dramatic examples of habitat loss come from tropical rain forests. Once covering more than 14 per cent of the earth’s land surface, these rain forests now cover no more than 6 per cent. They are being destroyed fast. By the time you finish reading this chapter, 1000 more hectares of rain forest would have been lost. The Amazon rain forest (it is so huge that it is called the ‘lungs of the planet’) harbouring probably millions of species is being cut and cleared for cultivating soya beans or for conversion to grasslands for raising beef cattle. Besides total loss, the degradation of many habitats by pollution also threatens the survival of many species. When large habitats are broken up into small fragments due to various human activities, mammals and birds requiring large territories and certain animals with migratory habits are badly affected, leading to population declines.
(ii) Over-exploitation: Humans have always depended on nature for food and shelter, but when ‘need’ turns to ‘greed’, it leads to over-exploitation of natural resources. Many species extinctions in the last 500 years (Steller’s sea cow, passenger pigeon) were due to overexploitation by humans. Presently many marine fish populations around the world are over harvested, endangering the continued existence of some commercially important species.
(iii) Alien species invasions: When alien species are introduced unintentionally or deliberately for whatever purpose, some of them turn invasive, and cause decline or extinction of indigenous species. The Nile perch introduced into Lake Victoria in east Africa led eventually to the extinction of an ecologically unique assemblage of more than 200 species of cichlid fish in the lake. You must be familiar with the environmental damage caused and threat posed to our native species by invasive weed species like carrot grass (Parthenium), Lantana and water hyacinth (Eicchornia). The recent illegal introduction of the African catfish Clarias gariepinus for aquaculture purposes is posing a threat to the indigenous catfishes in our rivers.
(iv) Co-extinctions: When a species becomes extinct, the plant and animal species associated with it in an obligatory way also become extinct. When a host fish species becomes extinct, its unique assemblage of parasites also meets the same fate. Another example is the case of a coevolved plant-pollinator mutualism where extinction of one invariably leads to the extinction of the other.
13.2 Biodiversity Conservation
13.2.1 Why Should We Conserve Biodiversity?
There are many reasons, some obvious and others not so obvious, but all equally important. They can be grouped into three categories: narrowly utilitarian, broadly utilitarian, and ethical.
The narrowly utilitarian arguments for conserving biodiversity are obvious; humans derive countless direct economic benefits from nature- food (cereals, pulses, fruits), firewood, fibre, construction material, industrial products (tannins, lubricants, dyes, resins, perfumes ) and products of medicinal importance. More than 25 per cent of the drugs currently sold in the market worldwide are derived from plants and 25,000 species of plants contribute to the traditional medicines used by native peoples around the world. Nobody knows how many more medicinally useful plants there are in tropical rain forests waiting to be explored. With increasing resources put into ‘bioprospecting’ (exploring molecular, genetic and species-level diversity for products of economic importance), nations endowed with rich biodiversity can expect to reap enormous benefits.
The broadly utilitarian argument says that biodiversity plays a major role in many ecosystem services that nature provides. The fast-dwindling Amazon forest is estimated to produce, through photosynthesis, 20 per cent of the total oxygen in the earth’s atmosphere. Can we put an economic value on this service by nature? You can get some idea by finding out how much your neighborhood hospital spends on a cylinder of oxygen. Pollination (without which plants cannot give us fruits or seeds) is another service, ecosystems provide through pollinators layer – bees, bumblebees, birds and bats. What will be the costs of accomplishing pollination without help from natural pollinators? There are other intangible benefits – that we derive from nature–the aesthetic pleasures of walking through thick woods, watching spring flowers in full bloom or waking up to a bulbul’s song in the morning. Can we put a price tag on such things?
The ethical argument for conserving biodiversity relates to what we owe to millions of plant, animal and microbe species with whom we share this planet. Philosophically or spiritually, we need to realise that every species has an intrinsic value, even if it may not be of current or any economic value to us. We have a moral duty to care for their well-being and pass on our biological legacy in good order to future generations.
13.2.2 How do we conserve Biodiversity?
When we conserve and protect the whole ecosystem, its biodiversity at all levels is protected - we save the entire forest to save the tiger. This approach is called in situ (on site) conservation. However, when there are situations where an animal or plant is endangered or threatened (organisms facing a very high risk of extinction in the wild in the near future) and needs urgent measures to save it from extinction, ex situ (off site) conservation is the desirable approach.
In situ conservation– Faced with the conflict between development and conservation, many nations find it unrealistic and economically not feasible to conserve all their biological wealth. Invariably, the number of species waiting to be saved from extinction far exceeds the conservation resources available. On a global basis, this problem has been addressed by eminent conservationists. They identified for maximum protection certain ‘biodiversity hotspots’ regions with very high levels of species richness and high degree of endemism (that is, species confined to that region and not found anywhere else). Initially 25 biodiversity hotspots were identified but subsequently nine more have been added to the list, bringing the total number of biodiversity hotspots in the world to 34. These hotspots are also regions of accelerated habitat loss. Three of these hotspots – Western Ghats and Sri Lanka, Indo-Burma and Himalaya – cover our country’s exceptionally high biodiversity regions. Although all the biodiversity hotspots put together cover less than 2 per cent of the earth’s land area, the number of species they collectively harbour is extremely high and strict protection of these hotspots could reduce the ongoing mass extinctions by almost 30 per cent.
In India, ecologically unique and biodiversity-rich regions are legally protected as biosphere reserves, national parks and sanctuaries. India now has 14 biosphere reserves, 90 national parks and 448 wildlife sanctuaries. India has also a history of religious and cultural traditions that emphasised protection of nature. In many cultures, tracts of forest were set aside, and all the trees and wildlife within were venerated and given total protection. Such sacred groves are found in Khasi and Jaintia Hills in Meghalaya, Aravalli Hills of Rajasthan, Western Ghat regions of Karnataka and Maharashtra and the Sarguja, Chanda and Bastar areas of Madhya Pradesh. In Meghalaya, the sacred groves are the last refuges for a large number of rare and threatened plants.
Ex situ Conservation– In this approach, threatened animals and plants are taken out from their natural habitat and placed in special setting where they can be protected and given special care. Zoological parks, botanical gardens and wildlife safari parks serve this purpose. There are many animals that have become extinct in the wild but continue to be maintained in zoological parks. In recent years ex situ conservation has advanced beyond keeping threatened species in enclosures. Now gametes of threatened species can be preserved in viable and fertile condition for long periods using cryopreservation techniques, eggs can be fertilised in vitro, and plants can be propagated using tissue culture methods. Seeds of different genetic strains of commercially important plants can be kept for long periods in seed banks.
Biodiversity knows no political boundaries and its conservation is therefore a collective responsibility of all nations. The historic Convention on Biological Diversity (‘The Earth Summit’) held in Rio de Janeiro in 1992, called upon all nations to take appropriate measures for conservation of biodiversity and sustainable utilisation of its benefits. In a follow-up, the World Summit on Sustainable Development held in 2002 in Johannesburg, South Africa, 190 countries pledged their commitment to achieve by 2010, a significant reduction in the current rate of biodiversity loss at global, regional and local levels.
SUMMARY
Since life originated on earth nearly 3.8 billion years ago, there had been enormous diversification of life forms on earth. Biodiversity refers to the sum total of diversity that exists at all levels of biological organisation. Of particular importance is the diversity at genetic, species and ecosystem levels and conservation efforts are aimed at protecting diversity at all these levels.
More than 1.5 million species have been recorded in the world, but there might still be nearly 6 million species on earth waiting to be discovered and named. Of the named species, > 70 per cent are animals, of which 70 per cent are insects. The group Fungi has more species than all the vertebrate species combined. India, with about 45,000 species of plants and twice as many species of animals, is one of the 12 mega diversity countries of the world.
Species diversity on earth is not uniformly distributed but shows interesting patterns. It is generally highest in the tropics and decreases towards the poles. Important explanations for the species richness of the tropics are: Tropics had more evolutionary time; they provide a relatively constant environment and, they receive more solar energy which contributes to greater productivity. Species richness is also function of the area of a region; the species-area relationship is generally a rectangular hyperbolic function.
It is believed that communities with high diversity tend to be less variable, more productive and more resistant to biological invasions. Earth’s fossil history reveals incidence of mass extinctions in the past, but the present rates of extinction, largely attributed to human activities, are 100 to 1000 times higher. Nearly 700 species have become extinct in recent times and more than 15,500 species (of which > 650 are from India) currently face the threat of extinction. The causes of high extinction rates at present include habitat (particularly forests) loss and fragmentation, over-exploitation, biological invasions and co-extinctions.
Earth’s rich biodiversity is vital for the very survival of mankind. The reasons for conserving biodiversity are narrowly utilitarian, broadly utilitarian and ethical. Besides the direct benefits (food, fibre, firewood, pharmaceuticals, etc.), there are many indirect benefits we receive through ecosystem services such as pollination, pest control, climate moderation and flood control. We also have a moral responsibility to take good care of earth’s biodiversity and pass it on in good order to our next generation.
Biodiversity conservation may be in situ as well as ex situ. In in situ conservation, the endangered species are protected in their natural habitat so that the entire ecosystem is protected. Recently, 34 ‘biodiversity hotspots’ in the world have been proposed for intensive conservation efforts. Of these, three (Western Ghats-Sri Lanka, Himalaya and Indo-Burma) cover India’s rich biodiversity regions. Our country’s in situ conservation efforts are reflected in its 14 biosphere reserves, 90 national parks, > 450 wildlife sanctuaries and many sacred groves. Ex situ conservation methods include protective maintenance of threatened species in zoological parks and botanical gardens, in vitro fertilisation, tissue culture propagation and cryopreservation of gametes.
EXERCISES
1. Name the three important components of biodiversity.
2. How do ecologists estimate the total number of species present in the world?
3. Give three hypotheses for explaining why tropics show greatest levels of species richness.
4. What is the significance of the slope of regression in a species – area relationship?
5. What are the major causes of species losses in a geographical region?
6. How is biodiversity important for ecosystem functioning?
7. What are sacred groves? What is their role in conservation?
8. Among the ecosystem services are control of floods and soil erosion. How is this achieved by the biotic components of the ecosystem?
9. The species diversity of plants (22 per cent) is much less than that of animals (72 per cent). What could be the explanations to how animals achieved greater diversification?
10. Can you think of a situation where we deliberately want to make a species extinct? How would you justify it?